By: Lloyd Harris
What Happens In Colonies AFTER Mid August Determines How Successfully Colonies Can Be Wintered
If you ask most beekeepers how they prepare their colonies for winter, you will be told that “Winter colony preparation begins immediately after the current year’s honey has been removed.”
However, the exact calendar date will vary depending on the climate at the location where the bees are being kept.
A beekeeper’s primary objective at the end of the honey flow is to have colonies that are headed by a young “productive” queen and populated with a large population of healthy, well fed “young bees.”
Typically, Fall colony management begins with the beekeeper implementing their preferred method of Varroa mite control. This is usually followed by feeding colonies with enough sugar syrup to ensure that they do not starve during Winter.
During this process, colonies are usually checked to ensure that they contain a “productive “queen and populations are “adequate.” Then, the only thing left to do is to wait until ambient temperatures are cold enough before: moving the colonies to sheltered location and wrapping them with insulation; or moving them into overwintering quarters; or transporting them to a location where winter conditions are less extreme than those experienced on the Northern Great Plains of North America.
That is what the beekeeper is doing. The question is what are the bees doing? What is happening inside the colonies to enable them to survive Winter? When do colonies rear the bees that form the winter colonies and how many of them will still be alive come spring?
The answer to these questions depends on: 1) the colonies’ location, 2) the age of the queens, 3) the number of workers in the colonies, and 4) the beekeeper’s management style.
No matter where colonies reside, there is a seasonally related brood rearing pattern that controls how many bees are in each colony during Winter; although data showing these seasonal brood rearing patterns are scarce.*
Additional data sets about seasonally related brood rearing at other locations may exist in: proprietor research studies, unpublished graduate student thesis, or unpublished research projects. Many studies that reported measuring sealed brood during the season do not adhere to protocols that required sealed brood measurements be taken every 12 days (Smirl, 1970; Nelson, 1971; Fukuda, 1971). Even though, extensive data may have been collected during some of these research projects, if they were not attached to the original study they may now be unavailable or are lost.
In most cases, the seasonal brood rearing patterns in the available research projects are incomplete because they only covered part of the year as most of this research has been done by students and data collection ceased at the end of August when the researchers returned to classes.
In general, the colonies’ seasonal brood rearing patterns show increasing brood rearing levels beginning in spring, which continues until:
- the queen’s maximum egg laying capacity is reached sometime in July or August (Harris, 2008, 2010);
- honey storage encroaches on the area utilized for brood production (Harris, 2008; Geiger, 1967); or
- nectar or pollen availability declines (Smirl and Jay, 1972; Nelson and Jay, 1972; Mattila and Otis, 2007).
During this period, new worker bees are constantly being reared in colonies through the combined efforts of a colonies’ worker bees and their queens. How long these new bees exist in colonies is determined mainly by how long they normally live. This can change significantly if a colony:
becomes queenless (Fluri, 2012),
is moved to another apiary location (Nelson and Jay, 1989),
swarms, or
is exposed to insecticide(s).
Figure 1 shows the seasonally related worker bee brood rearing pattern and their resulting adult populations in the ten colonies located near Portage la Prairie, Manitoba on the Northern Great Plains of North America (Harris, 2008) during the Spring and Summer of 1975 and 1976. The differences in the sealed brood patterns between the two years reflected differences in local apiary conditions and the beekeeper’s managerial style.
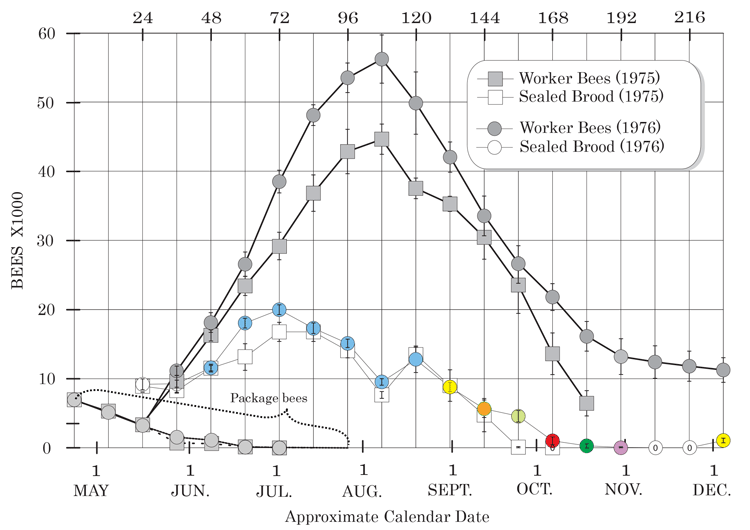
Figure 1. Average seasonal trends in adult worker bee populations and sealed brood in colonies started with 0.9 kg of worker bees in 1975 (n=5) and 1976 (n=5) in southern Manitoba, Canada. (Calculation Assumption: zero worker bee mortality in the first 12 days of adult worker bee’s life.
The 1976 sealed brood data after late August in Figure 1 has been colored to conform to the colored sub-populations represented in Figure 4. This sealed brood represents the portion of the sealed brood production curve that produced the early Winter colonies. Understanding what happens to the sealed brood production after the 31st of August is critical to understanding initial winter colony formation.
Figure 1 shows a steady decline in sealed brood production after mid-August, which continued until it temporarily ceased in October. The differences between the 1975 and 1976 sealed brood production curves after the honey flow shown in Figure 1 can be attributed to “a beekeeper effect”. Brood production responded to how the colonies were being managed.
The sealed brood data from September 1975 shows what happened when colonies did not receive any special late Summer-Autumn management. The consequences of leaving these colonies to prepare themselves for Winter were that these colonies:
- contained large quantities of canola honey, which they were unable to adequately utilize during Winter (see Figure 2),
- ceased brood rearing early (see figure 1),
- contained fewer bees (see figure 1),
- produced metabolic water that formed ice crystals on frames and on the inner cover above the bees (see Figure 3).
In 1976, the colonies were then managed to deal with these problems. The 1976 sealed brood data reflected what happened when colonies were fed sugar syrup in September and provided with an insulated inner cover on the 11th of November, before they were placed into a building in early December for Winter.

Figure 2. Crystallized canola honey residue unutilized by honey bees during Winter.
Although the general trends in sealed brood production after mid-August for the two years of data shown in Figure 1 followed the same general trends, there were apparent differences in brood production. These differences reflected changes in management practices.
In September 1976, the colonies were fed sugar syrup because it was apparent from the previous year that canola honey became too hard for the Winter bees to effectively utilize it during Winter (see Figure 2). Feeding sugar syrup to the colonies in September was only intended to provide colonies with a food source that would remain liquid longer. It not only did that, but it also appeared to have been responsible for the associated elevated sealed brood production that also occurred in September.

Figure 3. Ice formation on an uninsulated hive cover.
In early November 1976, colonies were also provided with insulated inner covers because of the amount of ice that had formed inside colonies from the metabolic water produced by the colonies the previous year (see Figure 3). The insulated inner covers appeared to have allowed the colonies to maintain conditions inside the hive that were conducive to brood rearing, which allowed colonies to resume brood rearing shortly thereafter. This was another unintended consequence of changing a management practice. Wrapping colonies with insulation for winter often produces a similarly effect and allows brood production to resume.
Although data showing brood rearing during late Summer, Autumn and Winter are scarce, this does not mean that brood rearing does not happen. It just means that researchers and beekeepers normally do not look inside their colonies once the honey flow is over and ambient temperatures fall below 320F (00C). However, brood rearing during Winter has been documented.*
It should be noted that low levels of brood rearing continued in these colonies throughout the Winter (Harris 2009), which is contrary to the commonly held belief that colonies suspend brood rearing during the Winter.
If a colony suspends its brood rearing during late Summer, Autumn or Winter, it usually indicates that it:
- contained an old queen,
- was queenless,
- was unable to maintain adequate brood rearing conditions, or
- contained insufficient stored pollen to support brood rearing.
If brood rearing does not continue in colonies from late Summer into Autumn or during Winter, the net result will be that their populations will continue to decline. There are no other options possible. Bees still die every day even during Autumn and Winter; albeit, at a much lower rate than those that occur during Spring and Summer (Nickel and Armbruster, 1937; El-Deeb, 1952; Free and Spencer-Booth, 1959; Sekiguchi and Sakagami, 1966; Mattila et al., 2001).
As of the 5th of December, only 32.3 percent of the adult bees from the 31st August sealed brood were still alive, only 67.4 percent of the bees from the 12th of September sealed brood were alive, only 74.2 per cent of the bees from the 24th of September sealed brood were alive, and 91.5 percent of the adult bees from the 6th of October sealed brood were still alive. Bees never stop dying. Without new bees being reared to replace the bees that are dying, the result was inevitable. The only thing that colonies could do was become smaller.
As the sealed brood production declined after the honey flow, so did the size of the adult worker bee population (Figure 1). Between mid to late August and the end of October, colonies declined at an average rate of approximately 481 bees per day, because these dying bees were not being replaced as quickly as they normally die. Consequently, adult bee populations declined rapidly after mid to late August.
The rapid population decline (see Figure 1) was not related to higher than normal worker bee mortality during this period. Worker bee longevity remained at levels similar to those normally observed in “healthy” colonies during Spring and Summer. Prior to the end of August, the average worker bee consistently lived between 33.3 to 36.0 days (Mattila et al, 2001) with only 0.085 percent of the adult worker bees living more than 84 days. Detectable changes in worker bee longevity did not occur until after early to mid-October when worker bees began living much longer than they did during Spring or Summer.
The reason for the rapid decline in the colonies adult bee populations during this period can be traced primarily to reduced brood rearing (see figure 1), which coincided with agricultural crops ceasing to flower. During this period, colonies simply did not rear enough new bees to compensate for the rate at which adult bees were dying.
Because all worker bees look virtually identical, it is almost impossible to tell which bees were added to a colony, when they were added, or how many from each age class still remain. There are ways to determine how old worker bees are, however all these methods are time consuming and involve intensive work.
The worker bee’s age can be determined by marking each new batch of emerging bees with a distinctive colored paint (Harris 1979) or tagging each bee with a small colored or numbered plastic disc (v. Ebert, 1922; Smith, 1972). These marked or tagged bees are then counted daily, weekly, or bi-weekly until they all die. It is then a matter of applying some basic mathematics to determine how many bees of what age remain in a colony (Harris, 1985; Harris, 2008; Harris, 2016). When the population estimates for each adult bee sub-population are arranged graphically in succession one above another, they illustrate the colony’s age structure or demography (see Figure 4).
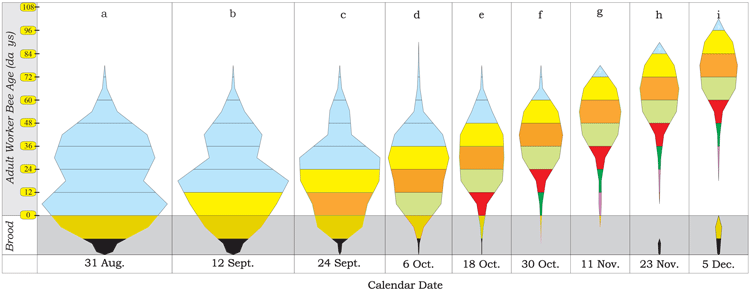
Figure 4 a-i.
Figure 4a illustrates a colony’s average age structure at the end of August/beginning of September for colonies started in Spring with two pound packages of worker bees and a mated queen. The unsealed brood (eggs & larvae) have been colored black. The sealed brood (sealed larvae and pupae) have been colored brown. All the adult bees that were present in the colonies on 31st of August were colored blue regardless of their age, because most of them, if not all of them, died before the 11th of November.
Colony age structures after the 31st of August are shown at 12 day intervals (Figure 4b-4i). Adult bee sub-populations emerging from the sealed brood after 31st of August are represented by a different color every 12th day. This allows the reader to easily follow each age related sub–population as they age and die.
Between the end of August and the 11th of November, all or almost all of the initial 42,038 (± 2,190) bees present in every colony at the end of August had died (refer to the blue colored bee subpopulation groups in Figure 4).
The “blue bees“ in Figures 4g, 4h, and 4i, were either the remnants of the adult bee sub-population that emerged as adult bees between the 19th of August and the 31st of August, or they are an artifact of the graphing methodology used to draw the graphs, which connected the mid-points of the respective sub-populations. Despite this subpopulation having been estimated to be zero on the 11th of November, there is a possibility that a small percentage of this sub-population may have been alive when the seasonal transition to longer lived Winter bees occurred.
The fate of all the “blue bee” sub-populations would still have been the same even if the initial colonies had been larger. These bees were all subject to the same Spring (or Summer) survival (or mortality) rates. Their life span was fixed. All of the bees present in the colonies at the beginning of December were reared between mid-August and late October.
In conclusion, what this reveals is that almost all the bees forming the Winter colonies in early December were produced from eggs laid after early August. In practical terms, this suggests that the colonies’ adult population’s size at the end of the honey flow may not be as important as one might have thought. Rather, it depends on when and how much brood is being reared and when the Autumn inflection in worker bee survival occurs (Mattila et al, 2001) that are important. The size of the colonies in early December is not directly related to the adult population’s size at the end of August. It depended entirely on how many bees colonies rear after mid-August. It may be that only a portion of the 42,038 (± 2,190) bees in every colonies on 31st of August were actually needed to rear the Winter colony.
The questions that beekeepers may want to ask themselves as they prepare their colonies for winter are:
What portion of the bees present in colonies at the end of the honey flow are actually needed to produce the Winter colonies or are these bees just part of an expensive retirement program; consuming valuable honey without contributing in any meaningful way to the Winter colony? It appears like the latter may be occurring.
What affect would re-queening colonies at the end of the honey flow have upon the production of the Winter colonies? At the end of the season, colonies with older queens cease brood rearing sooner than colonies with younger queens (Cale, 1956; Harris, 2008). It is likely that requeening colonies will alter their demographics or colony size, maybe both.
What will happen if management practices severely reduces brood rearing after mid-August, since this is when the early Winter colonies are produced? It appears that reduced brood rearing in August, September, or October will diminish how many bees colonies contain during Winter.
These three items need to be considered when colonies are being prepared for winter. What happens in honey bee colonies after mid-August determines how successfully the colonies can be wintered.
*The author has listed over 50 references supporting the information in this work. They have been only listed here in this article on Bee Culture’s web page for your review and can be accessed from the digital edition of this issue also.
References:
Allen, D. M. (1965) The Production of Queen Cups and Queen Cells in Relation to the General Development of Honeybee Colonies, and its Connection with Swarming and Supersedure. Journal of Apicultural Research. 4: 121 – 141.
Avitabile, A. (1978) Brood rearing in honey bee colonies from late autumn to early spring. Journal of Apicultural Research. 17: 69 – 73.
Bodenheimer, F. S.; Ben-Nerya, A. (1937) One year’s studies on the biology of the honeybee in Palestine. Annals of Applied Biology 24: 385 – 403.
Cale, G.H. (1956) In The Pink For Winter. American Bee Journal. 96: 397 – 400.
Desborough, J. G. (1853) On the duration of life in the queen, drone and worker of the honey bee. The Entomological Society of London. 2: 145 – 171.
- Ebert, O. R. (1922) Zur Massenentwicklung der Bienenvölker. Archiv für Bienenkunde. 4: 1 – 26.
El-Deeb, A. A. (1952) Longevity of some races of the honeybee (Apis mellifera L) Ph.D. Thesis, Illinois State University, Urbana, Illinois.
Farrar, C. L. (1931) A measure of some factors affecting the development of the honeybee colony. University of Massachusetts, Ph.D. Dissertation.
Farrar, C. L. (1963) The overwintering of productive colonies. The Hive and the Honey Bee. ed. R.A. Grout, Dadant and Sons. Hamilton, Illinois.
Free, J. B.; Spencer-Booth, Y. (1959) The longevity of worker bees (APIS MELLIFERA). Proceedings of the Royal Entomological Society of London. (A) 34: 141 – 149.
Fukuda, H. (1971) Improvement of Bodenheimer’s Method for Establishing Individual Number in Honeybee Colonies. Journal Of The Faculty Of Science Hokkaido University Series VI. Zoology. 18: 128 – 143.
Fukuda, H. (1983) The Relationship Between Work Efficiency And Population Size In A Colony. Researches On Population Ecology. 25:249 – 263.
Fluri, P. (2012) Regulation der Lebensdauer bei Arbeiterinnen der Honigbienen: Bedeutung der Brutpflege, des Nachwuchses und der Volksgrösse. Agroscope Liebefeld-Posieux Science Nr. 544.
Geiger, J. E. (1967) A Report On Size and Times Of Installing Package Bees At The Brandon Experimental Farm. Saskatchewan Department of Agriculture Bee Lines. 28: 5 – 8.
Gruszka, J.; Peer, D.; Trembly, A. (1990) The impact of Acarapis woodi on the development of package bee colonies. Saskatchewan Department of Agriculture Bee Lines. 89: 9 – 24.
Gong Y.F. (1980) Apiculture (in Chinese) cited by W. Shi (2001).
Harris, J. L. (1979) A rapid method for colour-marking single honeybees with fluorescent paint. Journal of Apicultural Research. 18: 201 – 203.
Harris, J. L. (1980) A Population Model and its Application to the Study of Honey Bee Colonies. MSc. Thesis, University of Manitoba, Winnipeg, Manitoba, Canada.
Harris, J. L. (1985) A model of honey bee colony population dynamics. Journal of Apicultural Research. 24: 228 – 236.
Harris, J. L. (2008) Development of honey bee colonies initiated from package bees on the Northern Great Plains of North America. Journal of Apicultural Research. 47: 141 – 150.
Harris, J. L. (2009) Development of honey bee colonies on the Northern Great Plains of North America during confinement to winter quarters. Journal of Apicultural Research and Bee World. 48: 8 – 90.
Harris, J. L. (2010) The effect of requeening in late July on honey bee colony development on the Northern Great Plains of North America after removal from an indoor winter storage facility. Journal of Apicultural Research. 49:159 – 169.
Harris, J. L. (2015) Productive Fall Colony Management – Colony Establishment or Honey Bee Colony Management after the Honey Flow. Saskatchewan Agricultural Demonstration of Practices and Technologies (ADOPT) Project 20130301 – 19 pgs.
Harris, J. L. (2016) Thinking Inside The Box: Colony Demography or What Shape Is Your Colony In? Bee Culture. 144: 27 – 33.
Imdorf, A.; Ruoff, K.; Fluri P (2008) Volksentwicklung bei der Honigbiene. ALP Forum. 68: 1–88.
Jay, S.C. (1974) Seasonal development of honeybee colonies started from package bees. Journal of Apicultural Research. 13:149 – 152.
Jeffree, E. P. (1956) Winter brood and pollen in honey bee colonies. Insectes Sociaux. 2: 417 – 422.
Kostarelou-Damianidou, M.; Thrasyvoulou, A.; Tselios, D.; Bladenopoulos, K. (1995) Brood and honey production of honey bee colonies requeened at various frequencies. Journal of Apicultural Research 34: 9 – 14.
Liebig G. (1996) Entwicklung von Bienenvölkern, In der Fressensäckern 10, D-74321 Bietigheim-Bissingen, Festschrift der Gesellschaft der Freunde der landesanstalt für Bienenkunde der Universität Hohenheim.
Mattila, H. R.; Harris, J. L.; Otis, G.W. (2001) Timing of production of winter bees in honey bee (Apis mellifera) colonies. Insect Sociaux. 48:88 – 93
Mattila, H. R.; Otis, G.W. (2007) Dwindling pollen resources trigger the transition to broodless populations of long-lived honeybees each autumn. Ecological Entomology. 32: 496 – 505.
Merrill, J. H. (1924a) Bees and Brood from May until frost American Bee Journal. 64: 231 – 232.
Merrill, J. H. (1924b) Sealed and Unsealed Brood. American Bee Journal. 64: 424 – 425.
Merrill, J.H. (1925) Relation of Broodrearing to Honey Production. American Bee Journal 65: 118 – 119.
Moeller, F. E. (1952) The Effects of Stock Lines upon the Honeybee Population-Production Relationship. Ph.D. Thesis, University of Wisconsin, Madison, Wisconsin.
Murilhas, A. M. (2002) Varroa destructor infestation impact on Apis mellifera crania capped worker brood production, bee population and honey storage in a Mediterranean climate. Apidologie. 33: 271 – 281.
Nelson, D. L. (1971) Population growth study of package bee colonies in Manitoba. MSc. Thesis, University of Manitoba, Winnipeg, Manitoba, Canada.
Nelson, D. L.; Jay, S. C. (1972) Population growth and honey yield studies of package bee colonies in Manitoba. II. Colonies initiated with four package sizes on one date. Manitoba Entomologist. 6: 17 – 22.
Nelson, D. L.; Jay, S. C. (1989) The effect of colony relocation on loss and disorientation of honeybee. Apidologie. 10: 245 – 250.
Nickel, K. H.; Armbruster, L. (1937) Vom Ledenslauf der Arbeitsbienen besonders auch bei Nosemaschäden. 1. Teil. Archiv für Bienenkunde. 18: 257 – 287.
Nolan, W. J. (1923) A Two Year Curve for a Single Colony of Bees. Journal of Economic Entomology. 16: 117 – 124.
Nolan, W. J. (1925) Brood-rearing cycle of the honeybee. Bulletin United States Department of Agriculture No. 1349.
Nolan, W. J. (1928) Seasonal brood-rearing activity of the Cyprian honey-bee. Journal of Economic Entomology 21: 393 – 403
Nolan, W. J. (1932) Development of package-bee colonies. Technical Bulletin United States Department of Agriculture No. 309.
Pankiw, P. 1968) The Influence Of Environmental Conditions On Brood Rearing, Build-up, And Honey Production Of Package Bee Colonies. The Canadian Entomologist. 100: 127 – 134
Peer, D.; Gruszka, J.; Trembly, A. (1987). Preliminary observations on the impact of Acarapis woodi on the development of package bee colonies. Saskatchewan Department of Agriculture Bee Lines. 81: 6 – 13.
Quimby, M. (1863) Mystery of beekeeping explained. C.M. Saxton, New York. – cited by C.L. Farrar (1960) in: Old and new ideas about wintering. American Bee Journal. 100(8): 306 – 310.
Sekiguchi, K.; Sakagami, S. F. (1966) Structure of foraging population and related problems in the honeybee, with considerations on the division of labour in bee colonies. Report No. 69. Hokkaido National Agricultural Experimental Station.
Shi, W. (2001) Genetic Variation and Colony Development of Honey Bees (Apis Mellifera) in Kenya. Volume 291 of Swedish Univ. of Agricultural Sciences: Agraria, Acta Universitatis Agriculturae Sueciae.
Smirl, C. B. (1970) A study of the development and performance of package bees in Manitoba. MSc. Thesis, University of Manitoba, Winnipeg, Manitoba, Canada.
Smirl, C. B.; Jay, S. C. (1972) Population growth and honey yield studies of package bee colonies in Manitoba. 1. Colonies initiated with two package sizes on three dates. Manitoba Entomologist 6: 9 – 16.
Smith, M. V. (1972) Marking bees and queens. Bee World. 53: 9 – 13.
Szabo, T. I. (1993) Brood rearing in outdoor wintered honey bee colonies. American Bee Journal. 133: 579 – 580.
Wille, H.; Gerig, L. (1976) Massenwechsel des Bienenvolkes. IV. Zusammenspiel der Eilegetätigkeit der Königin, der Bienenschlupfrate der Arbeiterinnen. Schweiz. Bienen-Ztg. 99, 16-25, 125-140, 245-257.
Winston, M. L. (1979).IntraColony Demography and Reproductive Rate of the Africanized Honeybee in South America. Behavioral Ecology and Sociobiology 45: 279 – 292.
Winston, M. L. (1980). Seasonal patterns of brood rearing and worker longevity in colonies of Africanized honeybees in South America. Journal of the Kansas Entomological Society. 53: 157 – 165.